|
|
|
DNA-Forschung
Ergebnis Mütterlich-Großväterlich,
Rodolfo Berti
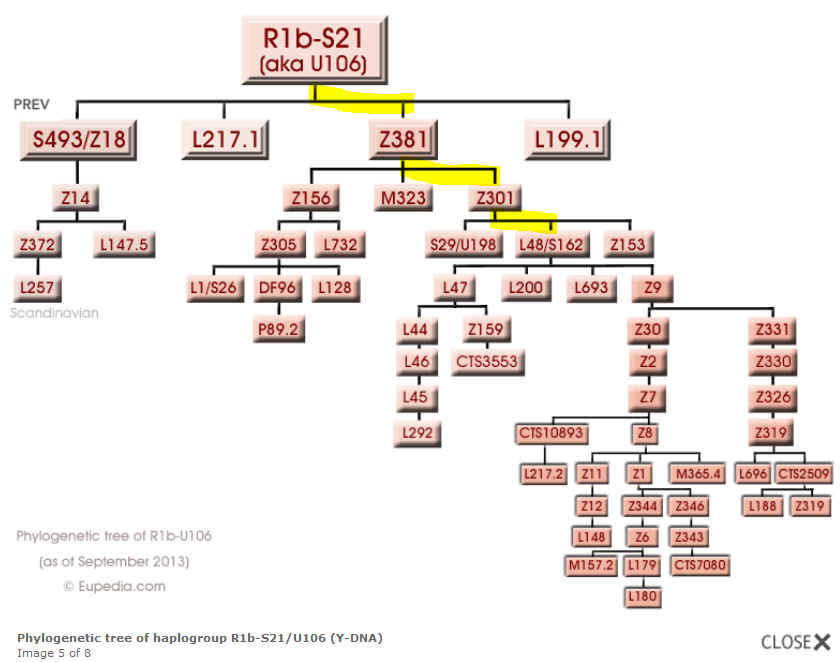
R 1b1a2a1a1a3a1a– M-269, U106 (Subglade
R-S26 (L-1))
R1b-S26 =>72% R1b-E.Europe =>15% R1b =>5% R1b-Ub =>5% R1b-C.Europe =>3%
Ergebnis Väterliche Linie,
Volker Wagner
R
1b1a2a1a1a3b2 – M269, U106, S162(Subglade R-S21 (L-48))
R1b1b2a1a1d
(nach 23andme)
R1b-Frisian3 =>38% R1b-S21* =>31% R1b-Frisian =>31%
Typisch
europäische Gentype, besonders stark verbreitet bei Bewohnern der
britischen Inseln, der Beneluxstaaten und Deutschland, aber auch in
Westfrankreich, Portugal und Galizien. Als Untergruppe R 1b1a2
außerhalb Europas kaum (Asien, Amerika, Naher Osten) bis gar keine
(Afrika) Verbreitung. Die Gruppe R1b wird vor allem mit den
Indogermanen in Verbindung gebracht. Gerne wird sie auch als die 'Kelten-DNA'
bezeichnet, da sie bei Völkern mit keltischen Hintergrund am
stärksten verbreitet ist, die Untergruppe U106 hingegen steht vor
allem für einen Teil der Germanen. Korrekter dürfte aber eine Verbindung
mit der steinzeitlichen Megalithkultur der Atlantikküste sein,
deren Vorfahren vor ca. 8000 Jahren einwanderten. Wahrscheinlich
lebten die Angehörigen von U106 im sogenannten Doggerland, welches
im Neolithikum und der Bronzezeit in mehreren Schüben von der
Nordsee überflutet wurde. Daher ist die Verbreitung dieser Menschen
vor allem entlang der deutschen Nordseeküste zu suchen, und
tatsächlich führt hier hin auch der Stammbaum, genaugenommen nach
Dithmarschen. Eine Friesische Herkunft meiner Ahnen ist hier
anzunehmen..
Herkunft
Es wird angenommen, dass die Haplogruppe R zwischen
Kaspischen Meer und Sibirien vor 30.000 bis 35.000 Jahren erstmals
auftrat. Die eher seltenen Formen der Chromosomen der Haplogruppe R,
sowie die häufigsten Fälle der mit ihr eng verwandten Haplogruppe
Q, werden interessanterweise unter Bevölkerungen in Mittelasien, in
Südasien, in Australien, in Sibirien, bei amerikanischen
Ureinwohnern, in Ägypten und in Kamerun gefunden. Sie ist die
häufigste Haplogruppe in Europa.
Einige Forscher wie Kivisild u.a. schlagen vor, dass
Süd- und Westasien der Ursprung dieser Haplogruppe" sein
könnte: Anhand der geographischen Verbreitung und der STR
Unterschiede der Schwesterngruppen R1 und R2 (letztere ist auf
Indien, Pakistan, Iran und das südliche Zentralasien beschränkt)
ist es möglich dass Süd- und Westasien die Quelle für die
Abgrenzung von R1 und R2 sind.
Verbreitung
Die Mehrheit der Träger von Haplogruppe R gehören
der Haplogruppe R1 an, die durch Marker M173 unterschieden wird. R1
ist in Europa und Westeurasien sehr verbreitet. Es wird angenommen,
dass seine Verbreitung mit der Wiederbesiedelung von Nord-Eurasien
nach der letzten Eiszeit zusammenhängt. Seine Hauptuntergruppen
sind R1a (SRY1532) und R1b (M343). Ein isolierter Stamm der
Y-Chromosomen, die scheinbar zu Haplogruppe R1b1* (P25) gehören,
wurde in hoher Konzentration unter der einheimischen Bevölkerung
von Nordkamerun in West-Zentralafrika gefunden. Es wird angenommen,
dass dies eine prähistorische Rückwanderung einer alten
protoeurasischen Bevölkerung nach Afrika darstellt. Einige Forscher
haben auch über Haplogruppe T Y-Chromosomen in geringer
Konzentration in einigen dieser kamerunischen Bevölkerungen
berichtet, die eine eurasische Ähnlichkeit aufweisen. Einige
Y-Chromosomen, die scheinbar mit den kamerunischen R1b1* Chromosomen
eng verwandt sind, werden in hoher Konzentration unter der modernen
Bevölkerung in Ägypten gefunden. Viele moderne Bevölkerungen des
Nordkamerun sprechen Tschadische Sprachen, die als ein alter Zweig
der Afro-asiatischen Sprachfamilie eingestuft werden. Die
ausgestorbene Sprache des Now der alten Ägypter gehörte auch der
gleichen Sprachfamilie an. Personen, deren Y-Chromosomen alle
Mutationen an den internen Nullpunkten des Y-DNA Baumes aufweisen
unter Einbeziehung von M207 (definiert Haplogruppe R), aber weder
die Mutation M173 (definiert Haplogruppe R1) noch die Mutation M124
(definiert Haplogruppe R2) besitzen, werden Haplogruppe R*
zugerechnet. Einige Fälle von Haplogruppe R* sind in den Proben der
australischen Aborigines gefunden worden. Haplogruppe R* wurde auch
in 10,3 % einer Probe von Burusho und in 6,8 % einer Probe von
Kalash in Nordpakistan entdeckt.
Ursprung R1b
Die Y-Haplogruppe R1b ist eine Abzweigung von R1
(M173), und wird durch den M343 Marker unterschieden. Der R1b Stamm
scheint einen viel höheren Grad an innerer Verschiedenartigkeit/Diversität
als R1a zu haben, was nahelegt, dass die Mutation M343, die R1b von
R1* trennt, beträchtlich früher aufgetreten sein muss als die
Mutation SRY1532, die R1a unterscheidet. Das Genographic Project
nimmt an, dass R1b auf der Iberischen Halbinsel als Refugium der
letzten Eiszeit entstanden ist, von wo aus sich die Gene danach
wieder ausbreiteten. Von dieser These wird inzwischen jedoch wieder
vermehrt Abstand genommen, da die Varianz (z.B. laut Barbara Arredi
und Kollegen) in Eurasien wesentlich höher ist und nach Westeuropa
hin kontinuierlich abnimmt, wobei jedoch gleichzeitig die Menge der
SNP zunimmt. Dies deutet darauf hin, das die iberische Population
wesentlich jünger ist. Dabei bildet auch die vermeintlich alte
baskische R1b-Population keine Ausnahme. Inzwischen geht man
vermehrt davon aus, das die Westeuropäer auf keinen Fall älter als
10.000 Jahre sind und vor diesem Zeitpunkt entweder im Maghreb oder
in vorderes Mittelasien lebten. Von Afrikanischen Wissenschaftlern
wurde die These geäußert das sich die Westeuropäer in der damals
noch fruchtbaren Sahara aufhielten und durch das Austrocknen der
Sahara über das Mittelmeer nach Südeuropa einwanderten. Dafür
spricht die Untergruppe V88 die eindeutig mit der Ausbreitung der
Tschadischen Sprachen (inkl. dem Altägyptischem) verbunden wird.
Zumindest im neuen Reich Ägyptens war R1b dort anwesend. Russische
Wissenschaftler sind dagegen der Meinung das sich R1b vom Altai aus
ausbreitete und ursprünglich eine Turk-Sprache nutzte. Sie
schließen R1b als Träger der Indogermanischen Ursprache aus.
Dieser These widerspricht jedoch die Existenz uralter R* und R1* im
Östlichen Iran, Pakistan, Afghanistan und Indien, wenn auch in
kleiner Menge, welche für eine Gemeinsamkeit zwischen R1b und
Indogermanen aufzeigt. Die Frage wie R1b also genau nach Europa kam,
ist daher nach wie vor ungelöst.
Die Haplogruppe R1b1a2 saß vor ca. 9.500 Jahren in
der Region um das Schwarze Meer. Die Einwanderung dieser Gruppe nach
Europa erfolgte frühestens mit der Ausbreitung der Landwirtschaft
ab 7.000 v.u.Z., höchstwahrscheinlich besteht auch ein starker
Zusammenhang mit den Indoeuropäern, die sich erst etwas später in
mehreren Wellen nach Europa ausbreiteten. In Ägypten macht diese
Gruppe heute unter 1% aus und ist sicher teilweise auf europäische
Einwanderung in der Bronzezeit zurück zu führen.
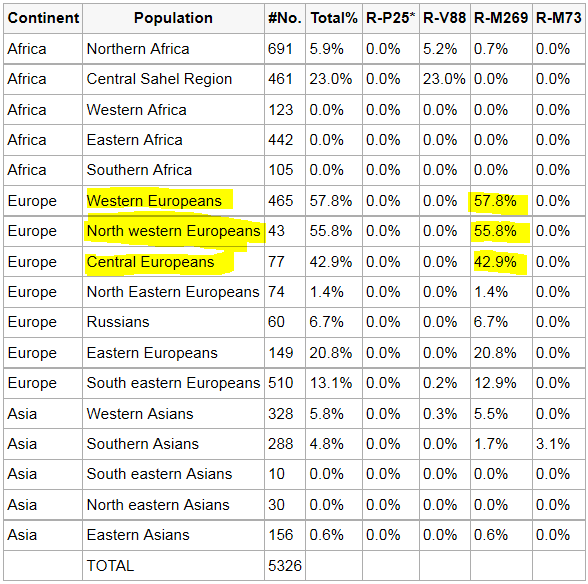
Heutige Verbreitung
In Europa ist R1b (mit den Untergruppen R1b1 und
R1b3, früher Hg1 und Eu18 benannt) die häufigste Y-Haplogruppe.
Die Konzentration erreicht in Teilen des nordwestlichen Irland 98 %;
in Nord- und Westengland, Spanien, Portugal und Irland bis 90 % und
im südöstlichen England und den Niederlanden noch etwa 70 %.
Darüber hinaus ist R1b in einigen Teilen Algeriens mit ca. 10 %
vertreten.
Berühmte Verwandte
Ein bekanntes Mitglied dieser DNA-Gruppe R1b1a2 ist
der ägyptische Pharao Tutanchamun. Tutanchamun war der letzte
Pharao der 18. Dynastie und regierte von 1332 bis 1323 v.u.Z.. Seine
väterliche Linie beginnt mit Pharao Thutmosis I. welcher etwa von
1504-1492 v.u.Z regierte. Dessen väterliche Herkunft ist unbekannt,
es ist deshalb derzeit noch unklar, wie diese Linie aus dem
Ursprungsgebiet nach Ägypten kam. Möglicherweise war sein Vater
Hethiter, einer indoeuropäischen Gruppe welche immer wieder gerne
in Verbindung mit Germanen gebracht wird und in der Antike in
Kleinasien ein mächtiges Reich gründeten. Aus der Regierungszeit
Echnatons bzw. Tutanchamuns ist aus den Archiven der Hethiter ein
Brief einer ägyptischen Königin überliefert. In diesem Brief
bittet sie den König der Hethiter um einen seiner Söhne als neuen
Pharao, da ihr Gatte tot sei und sie keinen Sohn habe. Die
Haplogruppe R1b1a2 war sicherlich auch im kleinasiatischen
Hethiter-Reich weit verbreitet. (Mein Gencode entspricht bei 16
Submarkern zu 50% dem Tutanchamun, auf 5 Markern gibt es eine
Abweichung von 1 Wert, auf drei Markern von zwei Werten. Hier
dürfte der gemeinsame Urahn vor ca. 4000-6000 Jahren gelebt haben.)
Verteilung Subclades R1
R1 ( M173 ) steht für die Entstehung in
Zentralasien
R1 ( L62 ) steht für eine Bevölkerung
die in den Ausläufern der anatolisch-iranischen Bergland lebt
R1 ( L62 *) sehr kleine Minderheit im
anatolisch-iranischen Bergland
R1 ( L120 ) prominenteste Subgruppe
innerhalb von L62
R1 ( L120 *) eine kleine Minderheit von
entfernt verwandten Linien in Anatolien und dem Kaukasus
R1 ( M17 ) steht für den wichtigsten
Teil (95%) in der L62-Gruppe
R1 ( L417 ) steht für 90% der L62, als
Untergruppe von M17
R1 ( Z645 ) steht bevorzugt für
Balto-Slawen in Europa und Indo-Iranern in Asien
R1 ( Z283 ) fast ausschließlich in
Europa, insbesondere mit baltischen und slawischen
Bevölkerungsanteil
R1 ( M458 ) steht für eine klar
Balto-slawische Abstammung
R1 ( L260 ) steht für eine weitgehend
westslawischen Gruppe
R1 ( Z280 ) steht für Balto-slawische
Abstammung, die mit ( M458 ) in Polen stark ist
R1 ( L365 ) wird vor allem in Polen
gefunden
R1 ( Z284 ) entstand in Norwegen,
verbreitet von den Wikingern
R1 ( Z93 ) spielt Hauptrolle in der
Verbreitung der indo-iranischen Sprachen
R1 ( M434 ) vor allem in Pakistan
R1 ( L664 ) stellt eine neolithische
Expansion in die deutschen Tieflandregionen
R1 ( M343 ) westliche Clade von R1 (
M173 ), mit Anteil asiatischer Herkunft ( L62 ) und steht für deren
westliche Ausdehnung
R1 ( P25 ) 5000 Jahre alte Mutation mit
Beschränkung von M343 R1b auf eine einzige Linie
R1 ( P297 ) erfolgreichste Linie in R1b,
möglicherweise im nordkaspischen Raum entstanden
R1 ( M73) heute als Minderheit in
Zentralasien, bei den Turkstämme und den pakistanischen Hazara
R1 ( M269 ) dominante Bevölkerung,
etabliert sich im Mesolithikum in der heutigen Ukraine
R1 ( L23 ) für eine Gruppe die die
pontische Steppe in Europa in der Jungsteinzeit querte
R1 ( L150 ) Gruppe welche die Mündung
der Donau erreichte, eine Mischpopulation von neolithischen Bauern
und altsteinzeitlichen Europiden
R1 ( L51 ) R1b drang im Neolithikum tief
nach Europa
R1 ( P310 ) steht für R1 ( M173 ) und
die Eroberung des europäischen Kernland um 2500 v. Chr. durch
eurasische Gruppen
R1 ( U106 ) trennt sich von der
Hauptgruppe ( P310 ) und absorbiert in der Altsteinzeit Europide
während ihres Auftreten in Deutschland
R1 ( Z18 ) westlichste Subgruppe von
U106, gilt als Ursprung der Germanen
R1 ( Z381 ) wichtigste Untergruppe von
U106, gilt ebenfalls als Ursprung der Germanen
R1 ( M467 ) Angelsächsischer Marker auf den
britischen Inseln und Norddeutschland
R1 ( L1) westgermanischer Marker auf den
britischen Inseln und Westeuropa
R1 ( L48 ) germanischer Marker auf den
britischen Inseln
R1 ( P312 ) zeigt primär
indoeuropäischen Bevölkerung seit der Bronzezeit Europa, vor allem
Kelten
R1 ( M65) steht für Ibero-keltischer
Abstammung.
R1 ( Z196 ) Wichtigste Ibero-keltischer
Abstammung beeinflusst die nordische Bronzezeit
R1 ( M153 ) Ebenfalls Ibero-keltischer
Abstammung, auch bei Basken gefunden
R1 ( L176.2 ) iberische Bevölkerung,
beteiligte sich an der nordischen Bronzezeit
R1 ( M167 ) ansehnliche Minderheit bei
Katalonen und Basken
R1 ( L165 ) steht für eine
Ibero-keltische Expansion nach Skandinavien während der nordischen
Bronzezeit
R1 ( U152 ) gallisch-keltische
Abstammung, Richtung Italien
R1 ( L21 ) britisch- keltische
Abstammung, vorherrschend auf den britischen Inseln
R1 ( M37 ) kleine Linie mit keltischen
Wurzeln
R1 ( M222 ) keltische Linie in Irland
R1 ( L513 ) Pan-britische keltische
Linie
R1 ( L96 ) keltische Abstammung, die
möglicherweise in Nordwest-Frankreich entstand
R1 ( L144 ) keltische Abstammung in
Irland
R1 ( L159.2 ) keltisch-britische Linie,
wurde sprachlich von den Angelsachsen unterjocht
R1 ( Z253 ) keltischer Abstammung in
Irland
R1 ( DF21 ) Pan-britische keltische
Abstammung
R1 ( L371 ) keltische Abstammung in
Wales
R1 ( L238 ) Nord-keltischen Abstammung,
spielte Rolle in der nordischen Bronzezeit gespielt
R1 ( DF19 ) Deutsch-keltische
Abstammung, teilt den gleichen Lebensraum mit der germanischen (
U106 )
R1 ( L584 ) anatolischer Zweig, der bei
Armenier, Assyrer und anderen anatolischen Bevölkerungen zu finden
ist
R1 ( M335 ) selten, Reste einer Linie
der Altsteinzeit in Anatolien
R1 ( V88 ) diese Mutation definiert alle
R1b Linien, die eine R1b Vorfahren in Afrika hatten
R1 ( V88 *) Cluster ist in Nord-Kamerun
R1 ( M18 ) Diese Bevölkerung wanderte
von Afrika südlich der Sahara in die Levante, und kam mit den
Phöniziern nach Sardinien
R1 ( V69 ) in der Jungsteinzeit hinter
der V88, am häufigsten in Nordkamerun
|
|
|
|
|
|
|
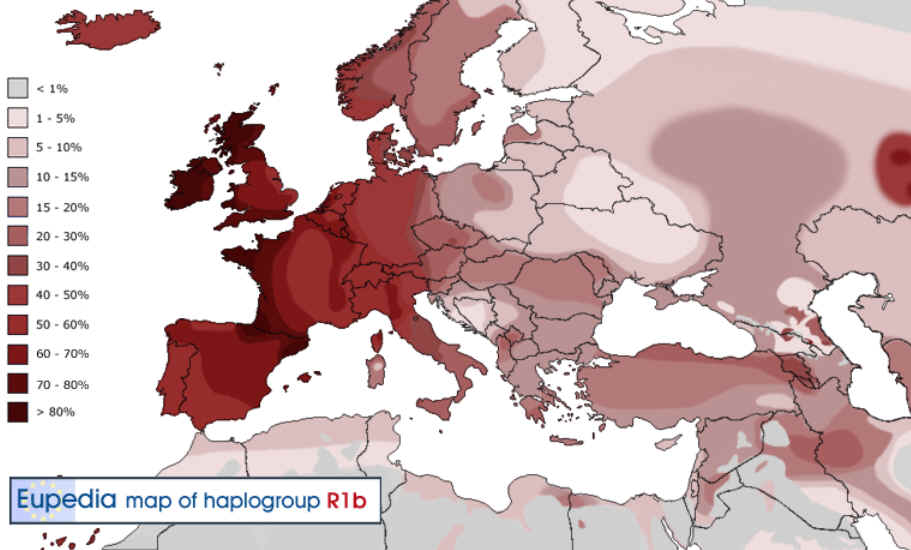
Verbreitung R1b in Europa |
|
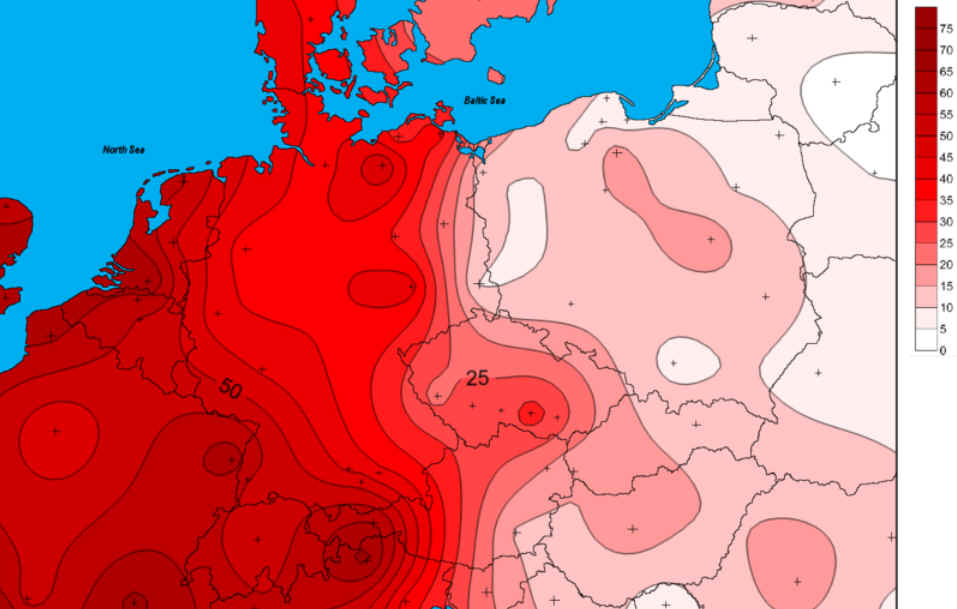
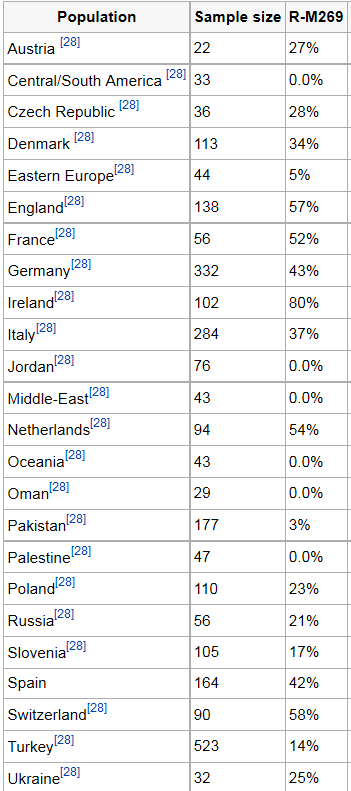
R1b in Mitteleuropa |
|
|
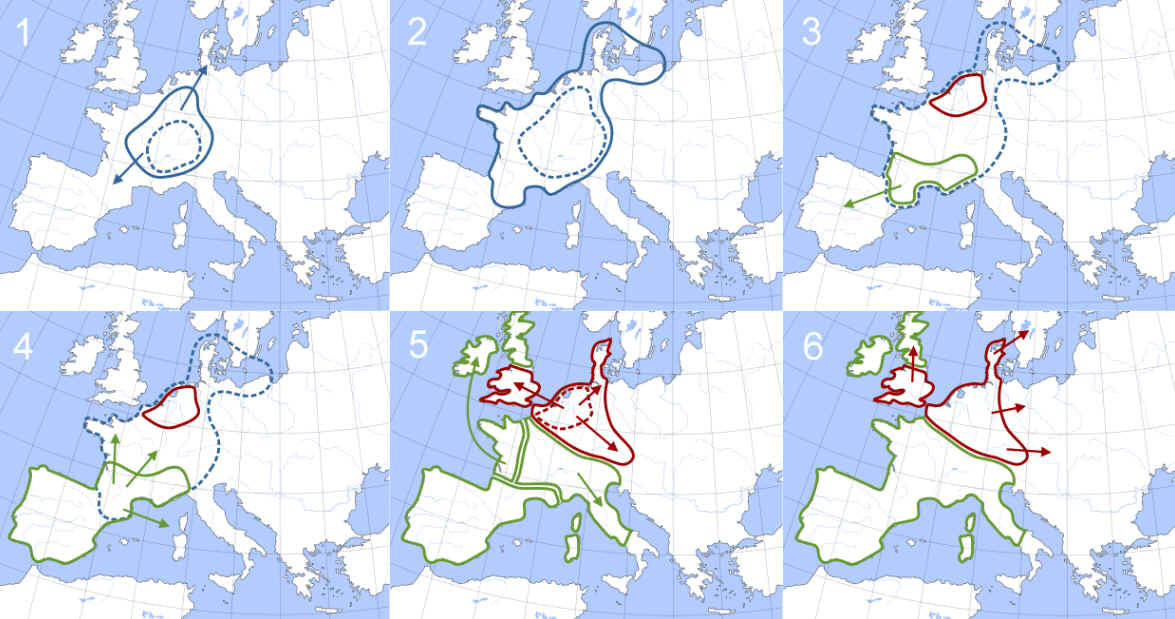
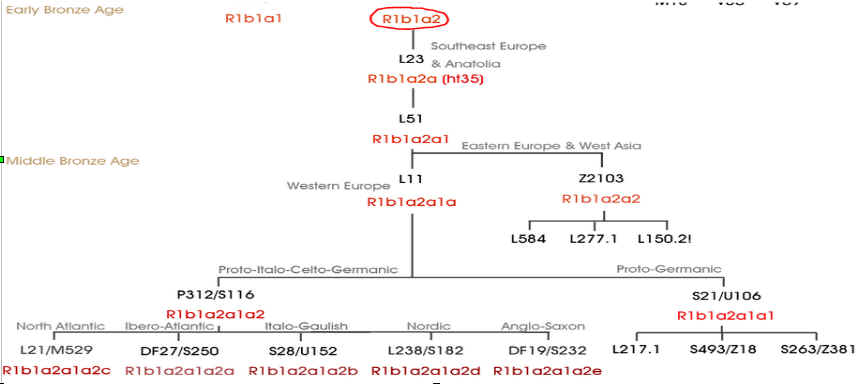
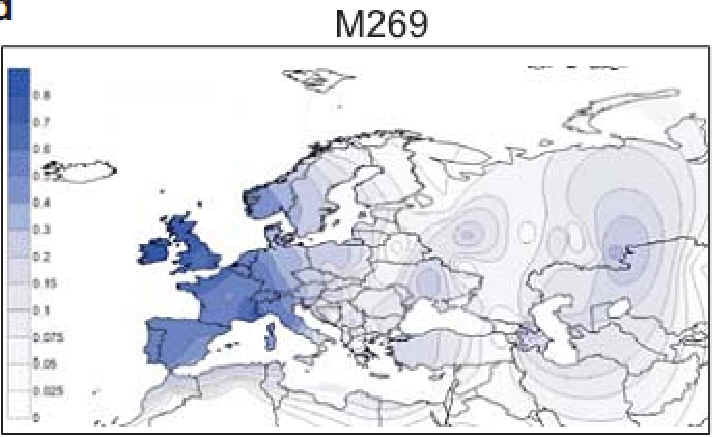
Entstehung R1b (blau) bzw. R1b-1a2
(Karmin) nach der Megalitherbauer-Theorie |
|
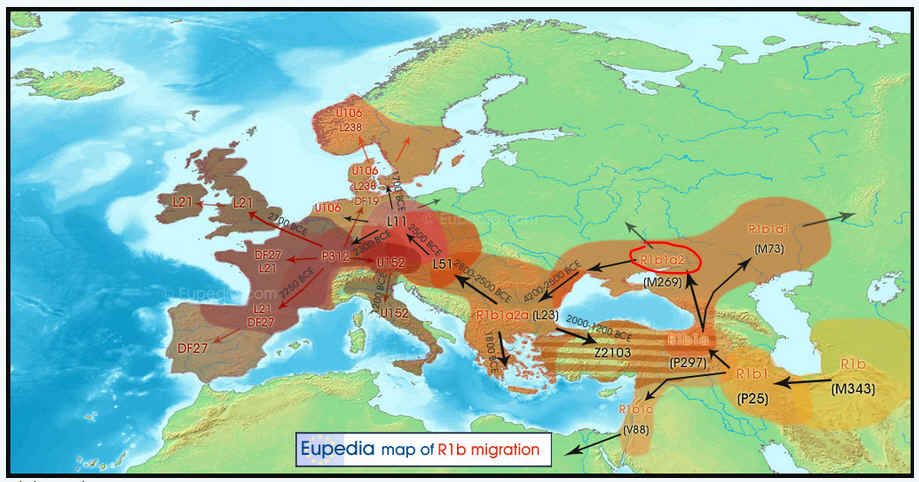
Entstehung und Verbreitung R1b / R1b1a2 nach der
Indogermanen-Theorie |
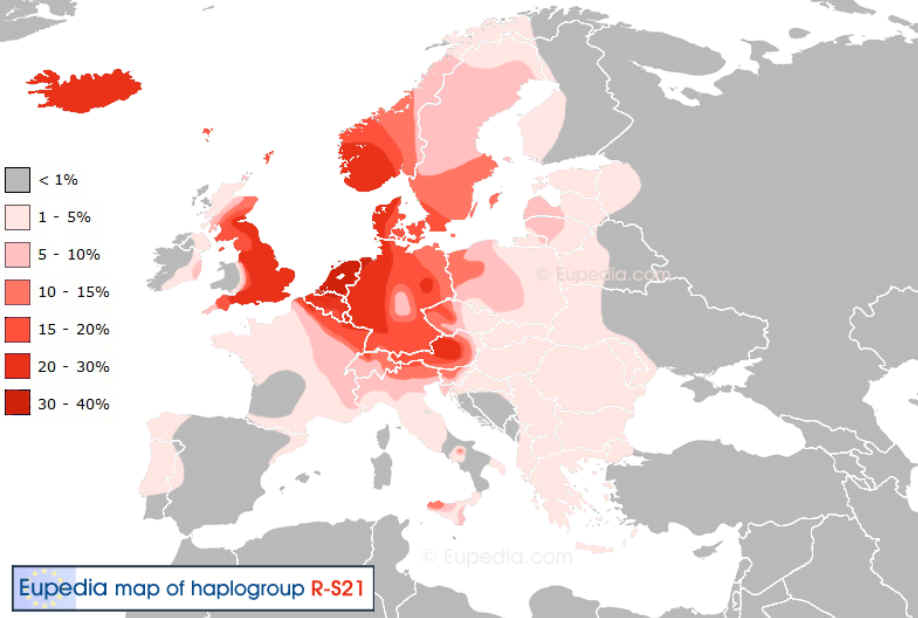
Verbreitung der eigenen Subglade |
|
|
|